We study sensory ion channels – the ion channels that serve as receptors for sensory stimuli such as acids, temperature and pain or are activated downstream of G protein-coupled receptor signaling
A New Family of Proton Channels: Identification of OTOP1 as a proton channel in taste receptor cells
The five basic tastes are detected by distinct subsets of taste receptor cells on the tongue and palate epithelium. Receptors for bitter, sweet and umami have been identified, but the receptor for sour remains elusive. To understand how the cells detect sour tastes, we first characterized the electrical response of taste cell from mice genetically modified to express a fluorescent reporter in their sour taste cells. When exposed to acidic solutions, we found that sour taste cells conducted an inward current carried by H+ ions that enter the cell (Chang et al, 2010). This H+ current is mediated by a novel ion channel.
We next set out to identify the gene that encodes this novel proton channel. Using transcriptome profiling (RNAseq), we identified genes whose expression was enriched in sour taste cells and that encoded unknown membrane proteins. Each candidate was then tested to determine if it generated a proton conductance when expressed in Xenopus oocytes or HEK-293 cells. Of the candidates tested, one, Otopetrin-1 (Otop1), encoding a protein with 12 transmembrane domains, generated a novel proton-selective channel able to conduct large H+ currents. Thus we conclude that Otop1 encodes a novel proton channel that is enriched in sour taste cells and is a candidate sour taste receptor (Tu et al, 2018).
We also discovered that homologs of Otop1 (Otop2 and Otop3) and invertebrate orthologs generate proton channels, each with distinct properties. These genes are expressed in unique distributions throughout the body, including in the digestive tract and in adipose tissue where their functions are not known.
For more information, see Tu et al, Science, 2018

OTOP1 functions as a sour receptor in mice
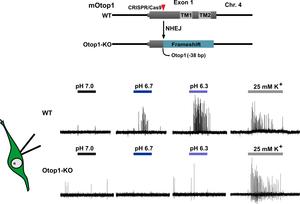
The identification of OTOP1 as a proton channel in sour taste cells (Tu et al, Science, 2018), strongly suggested that it funcitoned as the sour taste receptor. To test this directed, we generated a mouse strain in which the gene was inactivated by CRISPR/Cas9. In these OTOP1 knockout (KO) animals, we found that proton currents in sour taste receptor cells were completely eliminated. We next tested the effect of the Otop1 gene inactivation on sensory responses to acid stimuli in isolated cells and intact animals. Action potentials and changes in intracellular pH in response to acid stimuli were completely eliminated in taste receptor cells from KO animals, while gustatory nerves resposnes were strongly attenuated. Interestingly, despite the strong impact of the gene inactivation on sensory resposnes, Otop1 KO animals still avoided acids in two-bottle taste preference tests. Thus we demonstrated that OTOP1 is essential for gustatory responses to acid stimuli. This is strong evidence that OTOP1 functions as a sour taste receptor. How taste impacts animals behavior remains an open question and one we are are actively pursuing.
For more information, See Teng et al, Curr Biol, 2019
Cryo-EM Structure of OTOP1 and OTOP3
To visualize the structure of the newly identified OTOP channels, we teamed up with Andrew Ward at TSRI. By screening multiple species variants, zebrafish OTOP1 and chicken OTOP3 were found suitable for structural analysis.
Several major insights can be gleaned from these studies. First, they show that OTOP1 (and OTOP3) assembles as a homodimer. The twelve transmembrane helices form two structurally similar domains: an N domain which comprises the first six transmembrane helices and a C domain comprising the remaining six helices. This leads to a pseudo-tetrameric structure, with the four pseudo-subunits adopting a barrel shaped fold surrounding a central cavity.
Unlike other ion channels, the central cavity in the OTOP channels is filled with lipids and cannot support ion transport. Instead, three possible permeation pathways were identified, each of which contains a water-filled entrance leading to a constriction, or hydrophobic plug.
Ongoing work in the lab in collaboration with the Ward lab is aimed at determining how ions permeate the OTOP channels answering the question, where is the pore?. Other experiments are aimed at determining if the OTOP channels are gated (and by what) and at identifying structural rearrangements that gate the channels.

OTOP1 is a Sensor for the Taste of Ammonium Chloride
The otopetrin proteins (OTOPs) constitute a family of evolutionarily conserved proton-selective ion channels unrelated to other channels or receptors. Otopetrin 1 (OTOP1) conducts large inward proton currents in response to extracellular acidification and functions as the receptor for sour taste. Taste cells that express OTOP1 also mediate responses to high salts, including ammonium chloride (NH4Cl) which alkalinizes the cell cytosol, creating a driving force for proton entry. We, therefore, hypothesized that OTOP1 might function as a sensor for NH4Cl in the vertebrate taste system. To test this hypothesis, we expressed murine OTOP1in HEK-293 cells and measured currents with whole-cell patch clamp recording. We observed dose-dependent inward currents in response to extracellular NH4+ . We simultaneously measured changes in intracellular pH with the fluorescent indicator pHlourin and observed a positive correlation between the NH4Cl-induced current magnitude and the degree of intracellular alkalization. Similar responses were observed for human OTOP1, whereas relative NH4Cl sensitivity was diminished in zebrafish OTOP1 and enhanced in chicken OTOP1. The large magnitude of the responses and species differences suggested that OTOP1 responses to NH4Cl might involve the gating of OTOP1 by intracellular alkalinization. To test this, we generated mutations of conserved basic residues on the intracellular surface of mOTOP1 and identified one mutation, R292A, located in the linker joining tm 6-tm 7 which selectively reduced ammonium sensitivity without affecting acid responses. We also showed, using an Otop1-/-mouse strain, that OTOP1 is required for sensory responses of taste cells to NH4Cl. Taste responses to NH4Cl measured from isolated Type III TRCs, or gustatory nerves were severely attenuated or eliminated in an Otop1-/- mouse strain. Behavioral aversion of mice to NH4Cl, retained in Skn-1a-/- mice lacking Type II TRCs, was entirely abolished in a double knockout with Otop1. These data together reveal an unexpected role for the proton channel OTOP1 in mediating a major component of the taste of NH4Cl and a previously undescribed channel regulation mechanism that is conserved across species. OTOP channels are expressed throughout the body and are likely to mediate responses to ammonia/ammonium under other contexts, in addition to taste. See Liang et al, 2023 for more details.

